| Mapa del sitio | Portada | Redacción | Colabora | Enlaces | Buscador | Correo |
|
|

|
| 15 março de 2007 |
Felipe A. P. L. Costa (*)
Um dos padrões biogeográficos conhecidos mais fundamentais é o aumento que ocorre na diversidade de espécies à medida que nos deslocamos dos pólos em direção ao equador. A variedade é maior nos trópicos, ao mesmo tempo em que as comunidades de zonas temperadas tendem a ser dominadas por um número reduzido de espécies. Há, por exemplo, muito mais espécies de árvores crescendo em um hectare de floresta tropical do que em um hectare de floresta temperada [1]. Resultados semelhantes têm sido obtidos por biólogos de campo trabalhando nos mais diferentes tipos de hábitats e envolvendo uma ampla variedade de grupos de organismos, de aves, borboletas e formigas a peixes de água doce e moluscos marinhos, passando por ervas miúdas e fungos que vivem no solo.
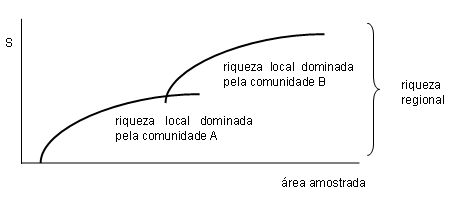
A expressão diversidade de espécies abrange, na realidade, duas grandezas distintas: riqueza e abundância relativa de espécies. A primeira diz respeito ao número de espécies que vivem em determinado hábitat, enquanto a segunda tem a ver com o tamanho de suas respectivas populações. Quando falamos estritamente em riqueza, estamos nos referindo apenas ao número de espécies presentes em determinado lugar, sem qualquer preocupação especial com a abundância relativa de suas populações. Duas ou mais comunidades podem abrigar números semelhantes de espécies, ainda que em proporções completamente diferentes. Um exemplo: sejam A e B duas comunidades, cada uma delas formada por 10 espécies de borboletas. Embora igualmente ricas, elas podem diferir bastante na abundância relativa das espécies que abrigam. Imaginemos uma situação extrema: na comunidade mais eqüitativa possível, todas as 10 espécies seriam igualmente abundantes (i.e., cada uma contribuiria com o mesmo percentual - no caso, 10 por cento - no cômputo dos indivíduos presentes); por usa vez, na comunidade menos eqüitativa possível, a grande maioria dos indivíduos (digamos, 91 por cento) pertenceria a uma única espécie dominante, restando uma parcela mínima (1 por cento, cada) para as demais espécies. Além de riqueza e abundância de espécies, comunidades também diferem em função da composição - i.e., a identidade das espécies (ou gêneros, famílias etc.) presentes em cada uma delas. Quer dizer, elas podem ser igualmente ricas e eqüitativas, mas ainda assim abrigar conjuntos distintos de espécies. Cada uma das comunidades de borboletas mencionadas no parágrafo anterior poderia ser formada por 10 espécies exclusivas. Nesse caso, nenhuma espécie encontrada em uma comunidade seria encontrada na outra e a soma das espécies encontradas nas duas comunidades duplicaria o número presente em cada uma delas em separado. Na maioria das vezes, porém, as comunidades partilham um número variável de espécies em comum, notadamente quando ocupam hábitats próximos e semelhantes. Há um amplo leque de possibilidades, mas podemos caracterizar a situação de comunidades particulares medindo o grau de similaridade entre elas. Comparar e classificar comunidades de acordo com a composição são preocupações antigas em ecologia e uma ampla variedade de técnicas e medidas - os chamados coeficientes de similaridade - já foi desenvolvida para isso. Uma razão para toda essa variedade são as sutilezas do mundo real. Um exemplo: sejam X e Y duas comunidades de tamanhos muito diferentes - 20 e 10 espécies, respectivamente. Se o tamanho das comunidades não é o mesmo, a parcela de espécies que há em comum entre elas também não pode ser a mesma. Vejamos como isso funciona: suponhamos que 10 por cento das espécies presentes na comunidade X também ocorram em Y. Todavia, como está última abriga apenas a metade do total de espécies da primeira, o percentual das espécies de Y que ocorrem em X seria outro (no caso, 20 por cento). A comparação entre duas comunidades pode dar origem, portanto, a dois valores de similaridade - da maior para a menor e da menor para a maior -, ao invés de apenas um. Mas as diferenças de riqueza estão longe de representar o maior desafio na comparação entre comunidades. Questões ainda mais problemáticas surgem quando a comparação leva em conta a abundância relativa das espécies presentes. Uma coisa é descobrir que duas comunidades têm 10 por cento de espécies em comum, todas ou quase todas elas sendo representadas por populações rarefeitas. Coisa bem diferente é descobrir que os mesmos 10 por cento em comum são formados por espécies com as populações mais abundantes. Há, mais uma vez, um amplo leque de combinações possíveis entre esses casos extremos. Questões adicionais, como o método de amostragem, devem ser levadas em conta pelo pesquisador na hora de escolher o coeficiente mais apropriado. Como dito antes, foi justamente para contornar essas dificuldades que diferentes coeficientes de similaridade foram desenvolvidos. Ainda assim, no entanto, todos eles apontam em uma mesma direção: quanto maior a semelhança na composição de espécies de duas comunidades, maior o grau de similaridade entre elas [2]. Em busca de estimativas confiáveis O leitor já deve ter percebido que, dependendo dos parâmetros utilizados (riqueza, abundância relativa, composição) na comparação entre comunidades, várias combinações de resultados são possíveis. Duas comunidades podem ser igualmente ricas e virtualmente idênticas em termos de composição, por exemplo, mas ainda assim diferir tremendamente na abundância relativa de suas espécies. Por causa disso, os índices usados para caracterizar comunidades de modo bastante resumido precisam incorporar mais de uma variável em suas fórmulas de cálculo. Esse é o caso do índice de Shannon-Weaver, talvez o índice de diversidade mais usado em ecologia, no qual riqueza e abundância relativa das espécies são combinadas de modo a produzir um valor numérico único para cada comunidade [3]. Outros índices de diversidade adotam procedimentos semelhantes. Nesse ponto, porém, precisamos ressaltar o seguinte: tanto riqueza como abundância relativa das espécies são variáveis essencialmente virtuais. O que isso significa? Significa que na grande maioria das vezes não é possível descobrir um valor exato para cada um desses parâmetros por meio de contagem direta. Na prática, temos de nos contentar com valores aproximados, obtidos por meio do emprego de alguma técnica de amostragem. Os valores assim obtidos representam estimativas do valor real da variável. Assim, trocamos uma pergunta aparentemente simples e direta - quantas espécies vivem aqui? -, por uma preocupação de natureza essencialmente metodológica: como obter estimativas fidedignas e confiáveis para a riqueza e a abundância relativas das espécies que vivem em determinado hábitat? Não estamos cutucando nenhum bicho-de-sete-cabeças, mas a resposta a esta pergunta não é de modo algum trivial. Razão pela qual, aliás, a questão tem sido debatida na literatura ecológica há mais de meio século [4]. Uma das dificuldades em responder a essa questão tem a ver com a falta de independência entre riqueza de espécies e número de indivíduos amostrados. Isso ocorre porque a riqueza costuma variar em função do tamanho da amostra (e.g., número de indivíduos amostrados) ou do esforço de amostragem (e.g., tempo gasto ou área coberta pelo observador). Para contornar problemas desse tipo, os estudiosos procuram calibrar suas amostras; um dos jeitos de fazer isso envolve a construção da chamada "curva do coletor". Mas como construir uma curva dessas? Para começar, precisamos de um par de eixos ortogonais. No eixo horizontal, colocamos a escala que mede o esforço de amostragem (e.g., quantidade de indivíduos ou área amostrada); no eixo vertical, colocamos a escala que mede a riqueza de espécies. Em seguida, admitindo que a parte mais trabalhosa (i.e., obter dados) foi devidamente equacionada e resolvida, distribuímos os valores obtidos no espaço criado por esse par de eixos ortogonais. Ao final do processo, se tudo funcionar direito, obteremos uma curva do coletor. Inúmeras curvas desse tipo já foram construídas por muitos pesquisadores trabalhando em diferentes tipos de hábitats, com os mais variados grupos de organismos. Apesar de toda essa variação, no entanto, há uma notável convergência no aspecto geral dessas curvas e as razões para que isso ocorra são relativamente fáceis de entender: primeiro, simplesmente porque no início do processo de amostragem a quantidade de espécies "novas" (não-capturadas) é bem maior do que a quantidade de espécies já capturadas. Por causa disso, a trajetória da curva do costuma ter um segmento inicial ascendente, indicando que nessa fase a amostragem de uma quantidade crescente de indivíduos (eixo horizontal) é traduzida em um número crescente de espécies (eixo vertical). Todavia, à medida que o trabalho de amostragem prossegue, a quantidade de espécies novas (não-amostradas) declina. Chega então um momento a partir do qual a maioria dos indivíduos amostrados passa a ser classificada simplesmente como "repetição" (i.e., indivíduos de espécies já amostradas antes). Assim, após uma fase inicial de crescimento acelerado, a curva prossegue cada mais devagar até se estabilizar em uma região de platô - a chamada assíntota da curva -, indicando que há um ponto a partir do qual a quantidade de espécies amostradas permanece mais ou menos inalterada, a despeito da continuação no processo de amostragem. Em termos metodológicos, o advento desse platô pode ser interpretado como o momento mais indicado para interromper os esforços de amostragem dentro daquele hábitat. Isso porque à medida que nos aproximamos do número total de espécies presentes (variável cujo valor real permanece virtualmente desconhecida), as chances de encontrar espécies novas tornam-se cada vez menores, ao mesmo tempo em que o esforço de amostragem torna-se, em certo sentido, cada vez mais improdutivo. O valor da projeção do platô no eixo vertical costuma ser adotado como a melhor estimativa da riqueza (ou diversidade) local de espécies, entendida aqui como a riqueza (ou diversidade) de espécies em determinado hábitat. Nada impede, porém, que os esforços de amostragem prossigam em direção a hábitats vizinhos. Quando isso ocorre, a trajetória do gráfico tende a retomar a trajetória ascendente descrita antes, embora isso agora ocorra a partir de um patamar inicial diferente de zero. Um exemplo do tipo de resultado que pode ser obtido com essas amostragens seqüenciais (no caso, envolvendo dois hábitats distintos), pode ser visto no gráfico abaixo:
Como dito antes, o eixo horizontal (área amostrada) representa o esforço de amostragem, enquanto o eixo vertical (S) indica a riqueza de espécies. Quando há resultados como esse disponíveis para uma série de comunidades locais (A, B etc.), torna-se possível então combinar os diferentes valores obtidos com o propósito de estimar a riqueza regional de espécies. Dependendo de como esse processo de amostragem é conduzido, podemos obter uma cadeia de resultados interligados (hierarquizados). Eis aqui um exemplo numérico, envolvendo valores reais da riqueza de espécies para um grupo de plantas que estudo: cerca de 35 espécies do gênero Solanum já foram encontradas no município de Juiz de Fora, 21 das quais na Reserva Biológica do Poço D'Anta. As espécies juizforanas são um subconjunto das 51 espécies registradas para a Zona da Mata mineira [5]; estas, por sua vez, integram a flora estadual, que contém 100-110 espécies do gênero, e estas, a flora brasileira, que abriga um total de aproximadamente 350-400 espécies. Estes números são provisórios e estão sujeitos a revisão, mas servem para ilustrar questões importantes que os estudiosos de comunidades estão procurando responder. Para início de conversa, podemos notar que há um aumento no número de espécies à medida que ampliamos o foco geográfico (ou "área de amostragem"); esse aumento, no entanto, é bem inferior ao que seria esperado se fizéssemos uma transposição direta de acordo com as dimensões da área sob consideração. Vejamos: levando em conta a densidade de espécies de Solanum na Reserva do Poço D'Anta (21 espécies em 2,77 quilômetros quadrados de área), poderíamos esperar números bem superiores aos mencionados, em todos os níveis hierárquicos (município, região estadual, estado, país). Para o município de Juiz de Fora (1,5 mil quilômetros quadrados), por exemplo, seriam esperadas mais de 11 mil espécies, e não as 35 registradas; para o estado de Minas Gerais (cerca de 588,4 mil quilômetros quadrados), seriam mais de 4,5 milhões de espécies, e não as 100-110 registradas. Qual a razão para tamanho disparate entre riqueza esperada e riqueza observada? A principal explicação tem a ver com o elevado grau de redundância que há na composição de comunidades locais de Solanum. E a razão por trás disso é que espécies com uma ampla área de distribuição geográfica terminam fazendo parte de inúmeras comunidades locais. A maioria das espécies registradas em Juiz de Fora, por exemplo, é ou já foi encontrada também em outros municípios da Zona da Mata mineira, várias delas em outras regiões do estado e algumas em outros estados brasileiros. Por fim, em que medida a composição de comunidades locais é influenciada pela disponibilidade regional de espécies? Seriam as 21 espécies de Solanum encontradas na Reserva do Poço D'Anta apenas uma amostra aleatória do conjunto de todas as espécies presentes no município de Juiz de Fora ou foram elas de algum modo reunidas ao longo do tempo por forças ecológicas organizadoras? Ainda não conheço as respostas, mas posso afirmar que questões intrigantes como essas ajudaram a converter o estudo da organização de comunidades em uma das áreas mais efervescentes da pesquisa científica contemporânea [6].
Notas
(*) Biólogo meiterer@hotmail.com, autor de ECOLOGIA, EVOLUÇÃO & O VALOR DAS PEQUENAS COISAS (2003, Edição do autor) e A CURVA DE KEELING E OUTROS PROCESSOS INVISÍVEIS QUE AFETAM A VIDA NA TERRA (2006, Editora Moderna).
1. Estamos tratando aqui de diferenças no número de espécies, não no número de árvores individuais. Quer dizer, um hectare de floresta temperada pode ter a mesma quantidade de árvores por unidade de área que uma floresta tropical; o que nos interessa aqui é que esta última tem uma variedade de árvores muito maior do que aquela. |
||