| Portada | Directorio | Buscador | Álbum | Redacción | Correo |
|
|

|
| 13 de julho de 2005 |
Felipe A. P. L. Costa (*)
A ecologia de insetos é uma disciplina científica desequilibrada: enquanto a maior parte dos estudos de campo tem sido conduzida em países de clima temperado, a grande maioria das espécies vive exclusivamente em hábitats tropicais. Além disso, como comumente estamos mais preocupados com insetos que podem causar danos ou estragos ao nosso bem-estar ou à economia, a literatura entomológica termina sendo uma amostra duplamente distorcida do mundo natural: há um número desproporcionalmente maior de estudos envolvendo pragas e espécies de zonas temperadas. A despeito de nossa reconhecida biodiversidade, o estudo de populações naturais de insetos - em contraste, por exemplo, com estudos de laboratório - ainda está dando seus primeiros passos entre nós. Este artigo relata um dos primeiros e, ainda hoje, um dos poucos estudos de campo conduzidos no país com o intuito de construir tabelas de vida, procurando com isso descrever o impacto relativo das fontes de mortalidade sobre uma população de insetos.
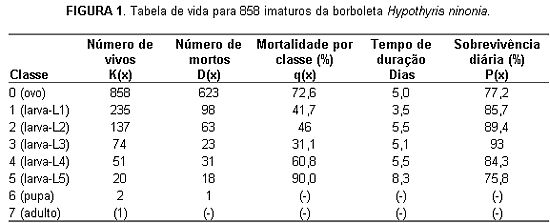
Populações biológicas são coleções de indivíduos de uma mesma espécie que vivem temporariamente juntos, em determinado lugar. Biólogos e outros cientistas que lidam com populações naturais comumente estão interessados em descrever e explicar a ocorrência e a magnitude das flutuações numéricas que elas exibem ao longo do tempo. O tamanho populacional é um balanço momentâneo entre ganhos (nascimentos e imigrações) e perdas (mortes e emigrações) de indivíduos: quando os ganhos superam as perdas, a população cresce; quando as perdas são maiores, ela diminui. Em muitas situações, no entanto, é bastante razoável admitir que a população esteja fechada (i.e., não saem nem chegam indivíduos); nesses casos, o tamanho é um balanço direto entre nascimentos e mortes e o estudo das fontes de mortalidade torna-se especialmente valioso para entendermos as flutuações populacionais. O impacto e a importância relativa das fontes de mortalidade que atuam sobre uma população podem ser avaliados por meio de uma técnica conhecida como análise de tabelas de vida [1]. Tabelas de vida procuram capturar e refletir as mudanças numéricas que ocorrem ao longo do tempo nos padrões de sobrevivência dos integrantes de uma população. Embora a construção de tabelas de vida para populações humanas tenha uma história relativamente antiga e uma relevância mais ou menos óbvia (políticas de seguridade social e a indústria do seguro de vida, por exemplo, dependem da análise de tabelas de vida), sua utilização em populações animais - e de insetos, em particular - é um empreendimento bem mais recente. São particularmente raros os estudos publicados envolvendo a construção de tabelas de vida para populações naturais de insetos que vivem em hábitats tropicais [2]. Essa carência não deixa de ser surpreendente, levando-se em conta que o tipo de trabalho de campo requerido não apresenta grandes dificuldades, exceto, talvez, pela necessidade de se fazer registros detalhados por períodos prolongados de tempo (um ano ou mais). Um modo de obter dados para construir uma tabela de vida é acompanhar o destino de uma coorte (conjunto de indivíduos nascidos em uma mesma data ou época do ano), desde o nascimento até a morte do último sobrevivente (tabela de vida horizontal). Isso não sendo possível, como muitas vezes não é, pode-se ainda tentar conduzir censos periódicos da população de interesse, classificando e contando os indivíduos encontrados em diferentes idades ou estágios do ciclo de vida (tabela de vida vertical). Nos insetos com metamorfose completa (o que inclui a maioria deles: besouros; mariposas e borboletas; vespas, abelhas e formigas; moscas e mosquitos etc.), o ciclo de vida é dividido em quatro estágios: ovo, larva, pupa e adulto. Entre as borboletas, mais especificamente, o estágio larval é ainda subdividido em cinco estádios larvais (L1 a L5), intercalados entre si por quatro períodos de muda larval e terminando com um período de muda pupal, quando larvas-L5 bem-sucedidas entram no estágio de pupa. O objetivo deste trabalho foi investigar os riscos de mortalidade dos imaturos (ovos e larvas) de uma população de borboletas, Hypothyris ninonia daeta (doravante referida como Hypothyris), na Reserva Biológica do Poço D'Anta, em Juiz de Fora (MG). Nessa reserva, fêmeas grávidas dessa borboleta depositam seus ovos (um ou dois de cada vez) na face inferior das folhas do braço-de-preguiça (Solanum cernuum) [3], onde as larvas permanecem se alimentando e, se conseguirem sobreviver, empupam. Ovos e larvas podem ser atacados e servir de alimento para um elenco variado de inimigos naturais, incluindo predadores (aranhas, besouros, carrapatos, formigas, marimbondos, percevejos e aves) e parasitóides (moscas e vespas). Entre janeiro de 1984 e junho de 1985, foram realizadas vistorias periódicas em um total de 96 braços-de-preguiça. Em cada planta vistoriada, eram registrados: a quantidade e localização dos ovos; e a quantidade, localização e tamanho (comprimento) das larvas. Como foi possível reconhecer ovos e larvas individuais, imaturos desaparecidos entre visitas sucessivas foram considerados como mortos e a proporção deles vista pela última vez em determinado estágio/estádio foi tratada como sendo igual à taxa de mortalidade naquele período. Por exemplo, se 40 ovos foram encontrados em uma visita, dos quais apenas 10 foram reencontrados na visita seguinte (como ovos ou larvas-L1), então a taxa de mortalidade dos ovos para o intervalo de tempo em questão foi estimada como sendo igual a (40 - 10) / 40 = 0,75 (ou 75 por cento). Chances de sobrevivência Ao final dos 18 meses de trabalho de campo, foram computados 878 imaturos de Hypothyris. Excluindo 20 ovos coletados ou danificados durante as vistorias, foram construídas tabelas de vida para os totais de imaturos encontrados em cada estação (verão de 1984 ao outono de 1985), envolvendo um total de 858 imaturos, dos quais (i) 623 (72,6%) morreram no estágio de ovo; (ii) 233 (27%) morreram em algum estádio larval; e (iii) dois conseguiram empupar (uma pupa deu origem a um adulto, a outra morreu parasitada). Não foram detectadas diferenças expressivas nas tabelas de vida sazonais; assim, por simplificação, apresento aqui uma tabela única para os 858 imaturos. A tabela da figura 1 mostra as classes que foram efetivamente investigadas: o estágio de ovo e os cincos estádios larvais. A raridade das pupas impediu que esse estágio fosse adequadamente incluído nas análises e discussões. Afinal, como essa tabela foi construída e o que exatamente os números apresentados significam?
Ao lado da coluna que mostra os estágios/estádios do ciclo de vida das borboletas Hypothyris, há uma primeira coluna numérica, K(x), contendo os dados fundamentais da tabela: o número de imaturos vivos no início de cada classe. O valor de K(1), por exemplo, indica o número de indivíduos vivos (235) no início do estádio de larva-L1. Todas as outras colunas são derivadas dessa coluna inicial. A coluna seguinte, D(x), mostra o número de indivíduos que morrem no transcurso de cada classe; o valor de D(1), por exemplo, indica o número de imaturos mortos no estádio de larva-L1, resultado de uma subtração: D(1) = K(1) - K(2) = 235 - 137 = 98 larvas. A noção de que o número de sobreviventes de uma coorte diminui à medida que avançamos no tempo é intuitiva e mais ou menos óbvia; afinal, cedo ou tarde, todos sucumbem. Todavia, a forma e a velocidade dessa diminuição variam tremendamente de uma espécie para outra e até entre populações de uma mesma espécie. Para detectar eventuais variações nas chances de sobrevivência dos imaturos, à medida que eles avançam de uma classe a outra, devemos analisar em seguida o conteúdo da coluna seguinte, q(x). Os valores dessa coluna indicam o percentual de mortes que ocorrem em cada classe, em relação ao número de indivíduos vivos no início da própria classe. Quer dizer, os valores dessa coluna estimam a força de mortalidade característica de cada classe, de acordo com a expressão: q(x) = 100 * D(x) / K(x). O valor de q(4), por exemplo, indica que cerca de 61 por cento (100 * 31 / 51 = 60,78) dos imaturos que iniciaram o estádio de larva-L4 morreram antes de terminá-lo. Em outras palavras, de cada 100 larvas que iniciaram o quarto estádio, cerca de 39 em média conseguiram terminá-lo, passando então para o quinto estádio larval. A rigor, para que os valores da coluna-q(x) possam ser usados de um jeito consistente em análises comparativas, precisamos acrescentar um termo de ajuste. Isso é necessário porque o tempo de duração, no caso dos imaturos de Hypothyris, não foi exatamente o mesmo para todas as classes investigadas, como mostra a coluna "Tempo de duração" [4]. As diferenças no tempo de duração das classes não invalidam os comentários feitos até aqui; ocorre que, sem uma padronização, seria difícil decidir se um valor mais alto de q(x) representaria de fato um aumento efetivo na taxa de mortalidade ou se seria simplesmente conseqüência do fato de uma classe ser mais demorada do que uma outra. Como um modo de padronização, converti os valores da coluna-q(x) em percentuais diários de sobrevivência, P(x), cujos resultados estão mostrados na última coluna da tabela [5].
Para facilitar a visualização, os valores das colunas q(x) e P(x) foram então transformados no gráfico da figura 2. Nesse gráfico, é possível observar que a trajetória descrita pelos valores de q(x) lembra o formato de um "U", indicando que a taxa de mortalidade foi alta no início (ovo) e no final (L4, L5) do período investigado, diminuindo acentuadamente nas classes intermediárias (L1, L2, L3). Já a trajetória descrita pelos valores de P(x) adquire um perfil menos oscilante, indicando que a probabilidade diária de sobrevivência não variou tanto assim de uma classe para outra. Esses resultados contrastam fortemente com o que seria esperado com base na literatura especializada, rica em estudos conduzidos em regiões temperadas [6]. Para os valores de q(x), por exemplo, deveríamos esperar uma trajetória em formato de "U" invertido: taxas de mortalidade elevadas nos primeiros estádios larvais, antecedidas por valores comparativamente mais baixos no estágio de ovo e sucedidas por valores igualmente mais baixos nos últimos estádios larvais. Nesse ponto, vale notar que a taxa de mortalidade - i.e., os valores da coluna q(x) - pode ser vista como o resultado de uma combinação entre dois parâmetros: a probabilidade diária de sobrevivência durante uma determinada classe e o tempo de duração dessa classe. Uma diminuição na primeira ou um aumento na segunda contribuem igualmente para elevar os riscos de morte. No caso dos imaturos de Hypothyris, parte da variação observada nos valores de q(x) tem a ver com diferenças no tempo de duração das classes e parte com diferenças nos riscos diários de morte. Vejamos: de ovo até o quinto estádio larval, as probabilidades diárias de sobrevivência oscilaram entre um mínimo de 75,8 por cento (L5) e um máximo de 93 por cento (L3). Os valores mais baixos foram registrados nas classes extremas: 77,2 por cento no estágio de ovo e 78,8 por cento, no estádio de larva-L5. Há, porém, bons motivos para concluir que esses percentuais tenham sido produzidos por processos distintos. Primeiro, em função do tempo de duração: o quinto estádio larval é o mais demorado de todos (tanto no verão como no inverno), enquanto o estágio de ovo é relativamente curto. Assim, a elevada taxa de mortalidade (90%) sofrida pelas larvas-L5 parece ter sido mais uma conseqüência do tempo de duração do estádio, enquanto a dos ovos (70%) refletiria mais sua baixa probabilidade diária de sobrevivência. Observações posteriores indicaram que (i) os ovos são atacados por um elenco ampliado de inimigos naturais - além de predadores (e.g., formigas e larvas de besouros), algumas moscas e vespas parasitóides parecem restringir seus ataques a essa fase do ciclo de vida; e (ii) aves e principalmente marimbondos são predadores vorazes, mas que concentram seus ataques nas larvas grandes (L5, principalmente). Trocando em miúdos, ovos e larvas-L5 são fases arriscadas e que sofrem perdas severas, mas por motivos diferentes: os primeiros porque são apreciados e atacados por muitos inimigos naturais, os segundos porque são apreciados por uns poucos predadores vorazes. (Além do que, há aqui um efeito de rarefação: como larvas-L5 são quase sempre muito menos numerosas do que os ovos, a perda de uma mesma quantidade de indivíduos tem um impacto relativo muito maior quando ocorre na classe mais rara.) Enquanto isso, os três primeiros estádios larvais sofreram as menores taxas de mortalidade, cujos valores oscilaram entre 31,1 por cento (L3) e 46 por cento (L2). A taxa mínima foi registrada para as larvas-L3 e, pelo jeito, isso não foi conseqüência de uma redução no tempo de duração do estádio, mas sim de um aumento expressivo na probabilidade diária de sobrevivência. Desconfio que as chances de sobrevivência atingem seu máximo nessa fase porque as larvas-L3 têm o tamanho "errado" para alguns dos predadores que atacam os estádios larvais vizinhos. Vale notar que larvas pequenas (L1 e L2) tendem a ser atacadas e mortas por um elenco próprio de inimigos naturais (como formigas e vespas pequenas), bem diferente do elenco que ataca as larvas maiores (L4 e L5), que inclui marimbondos e aves insetívoras. Nesse caso, então, poderia ocorrer o seguinte: larvas-L3 seriam grandes demais para os inimigos naturais que atacam larvas diminutas e, ao mesmo tempo, pequenas demais para chamar a atenção de predadores que caçam presas maiores. De um jeito ou de outro, respondendo à pergunta do título, uma fêmea adulta de Hypothyris, que vive de 4 a 8 meses (um recorde, entre as borboletas), precisa colocar centenas de ovos durante seu tempo de vida para ter alguma chance de substituir a si própria (e a um parceiro) na próxima geração! Controle biológico Estudos adicionais serão necessários para que possamos testar essa e outras hipóteses explicativas envolvendo a ecologia de borboletas Hypothyris, suas plantas-hospedeiras e seus inimigos naturais. Ademais, muito mais estudos de campo precisarão ser realizados até que tenhamos um quadro de referência minimamente consistente para descrever e entender melhor os padrões de mortalidade que ocorrem em populações de insetos tropicais. A questão aqui, evidentemente, não é saber se a trajetória em "U" ou em "U" invertido está certa ou errada. Estamos apenas diante de um quadro incompleto, constituído principalmente por estudos conduzidos em países temperados e cujo alcance é, por isso mesmo, limitado. Portanto, antes de formularmos conclusões mais gerais sobre a dinâmica de populações de insetos, precisaremos alargar a base de dados (incluindo mais estudos de campo conduzidos nos trópicos) sobre a qual nossas teorias possam ser construídas. Ocorre que, além de desdobramentos teóricos, o assunto implica em conseqüências práticas relevantes e, às vezes, urgentes. Vejamos o caso do controle biológico de pragas. Em comunidades naturais, erupções populacionais de insetos herbívoros são eventos relativamente raros e, quando ocorrem, costumam ser revertidas em pouco tempo pela ação de fatores como escassez de alimento ou atuação de inimigos naturais. O que poderia então ocorrer com essas populações se a disponibilidade de plantas-hospedeiras aumentasse, ao mesmo tempo em que seus inimigos naturais fossem removidos da comunidade? Pois é uma combinação duplamente favorável como essa - comida farta e escassez de inimigos - que muitos herbívoros encontram em áreas de cultivo, quando então se tornam "pragas" (expressão essencialmente antropocêntrica), prosperando com facilidade e rapidez, com efeitos potencialmente devastadores. Pragas agrícolas são combatidas e eventualmente mantidas em níveis economicamente aceitáveis por meio da aplicação de venenos químicos. Os efeitos colaterais da presença dessas substâncias, no entanto, costumam ser desastrosos, inclusive para a saúde humana. Nos últimos anos, uma parcela cada vez mais expressiva de consumidores tem percebido os malefícios que os venenos químicos podem trazer ao seu bem-estar. Por sua vez, a busca de alternativas tem gerado uma demanda crescente por produtos agrícolas livres de agrotóxicos (incluindo os chamados "alimentos orgânicos"), embora esses produtos ainda sejam relativamente raros. Com a elevação no nível de exigência dos consumidores, a aplicação massiva, descontrolada e irregular de venenos tende a ser substituída por um controle químico-biológico mais moderado, no qual o uso de substâncias químicas em menor escala seria suplementado pela liberação periódica de inimigos naturais - notadamente patógenos (vírus e bactérias) e parasitóides que atacam ovos e larvas jovens do inseto-alvo. Em outras situações, já é perfeitamente possível confiar na ação exclusiva de agentes biológicos, embora estudos e pesquisas adicionais precisem ser conduzidos. Se os resultados obtidos com Hypothyris podem de alguma forma ser generalizados, então as estratégias de combate que dão ênfase aos estágios iniciais do ciclo de vida da praga precisam ser repensadas. Ao contrário do que possa parecer, pode ser mais indicado, por exemplo, garantir a presença de inimigos que ataquem a praga em seus estágios mais avançados de desenvolvimento (larvas mais velhas, ao invés de ovos e larvas jovens), como fazem os predadores insetívoros mais generalistas (marimbondos, aves, lagartos etc.). Ocorre que, para manter populações locais desses predadores, teríamos de oferecer tipos suplementares de presa (afinal, eles são generalistas), além da praga que se deseja controlar. O modo mais simples, barato e duradouro de se fazer isso seria mudar a paisagem monótona das áreas de cultivo, incluindo nela fragmentos de hábitats ocupados por comunidades naturais. Mais do que reservatórios de pragas adicionais, fragmentos florestais podem abrigar inimigos-chaves que, de outro modo, não conseguiriam se estabelecer na região. No fim das contas, está na hora de substituirmos o "desflorestar para cultivar a terra" por um lema mais inteligente para nossa agricultura: "florestar para melhorar".
Notas
(*) Biólogo meiterer@hotmail.com, autor do livro ECOLOGIA, EVOLUÇÃO & O VALOR DAS PEQUENAS COISAS (2003). Os dados sobre imaturos de Hypothyris mencionados neste artigo foram obtidos enquanto o autor era aluno regular do Programa de Pós-graduação em Ecologia, da Universidade Estadual de Campinas (Campinas, SP); parte desse período, como bolsista da CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior).
1. Detalhes técnicos e exemplos detalhados podem ser encontrados em Southwood, T. R. E. 1978. Ecological methods, 2nd edition. London, Chapman. |
|||