| Mapa del sitio | Portada | Redacción | Colabora | Enlaces | Buscador | Correo |
|
|

|
| 21 de fevereiro de 2006 |
Felipe A. P. L. Costa (*)
Quase todos os seres vivos dependem de fontes externas de calor para manter a temperatura interna do corpo. Trata-se de um modo antigo, seguro e bastante econômico de viver. Ectotermia é o nome dado a esse estilo de vida e os organismos que o utilizam são chamados de ectotérmicos. Plantas, fungos, micróbios e a grande maioria dos animais são ectotérmicos. Uns poucos animais, no entanto, gastam parcelas expressivas do orçamento produzindo calor por vias metabólicas, mantendo assim a temperatura do corpo em patamares relativamente elevados. A produção interna de calor é chamada de endotermia e os animais que adotaram esse estilo de vida, notadamente aves e mamíferos, são chamados de endotérmicos [1].
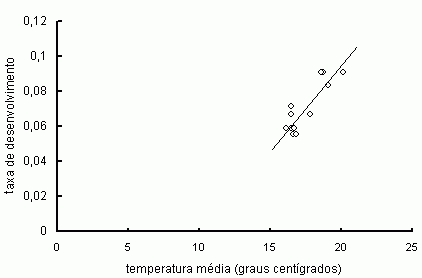
Aves e mamíferos também são às vezes chamados de animais de "sangue quente", uma alusão ao fato de que a temperatura do corpo deles permanece mais ou menos inalterada, a despeito de flutuações na temperatura externa. Quer dizer, mesmo quando a temperatura do ar cai muitos graus abaixo de zero, a temperatura corporal de um animal endotérmico permanece elevada - dando a impressão de que ele tem o sangue quente. Para conseguir isso, é necessário produzir calor, queimando boa parte dos materiais absorvidos durante a digestão. Uma vantagem evolutiva da endotermia pode ter sido justamente a de permitir que seus portadores mantivessem as atividades vitais em um patamar elevado, a despeito da temperatura ambiente [2]. Essa independência custa caro (basta dizer que boa parte do que comemos é gasto na produção metabólica de calor), mas tem também suas vantagens. É graças à endotermia, por exemplo, que as galinhas conseguem chocar seus ovos em 21 dias, a despeito da estação (fria ou quente) do ano. Mas aves e mamíferos são exceções. Para a grande maioria dos seres vivos, o ritmo de vida varia com as flutuações (diárias e sazonais) que ocorrem na temperatura do ar. Uma primeira explicação para isso é que o desempenho de atividades biológicas essenciais (respiração, fotossíntese, velocidade de locomoção etc.) varia em função da temperatura. Sendo assim, flutuações na temperatura deveriam ser acompanhadas por alterações nos índices de desempenho, especialmente no caso de organismos ectotérmicos [3]. Vamos explorar isso ao longo deste artigo, tendo como pano de fundo o estudo dos imaturos de uma borboleta. Lagartas em crescimento Lepidópteros (borboletas e mariposas) exibem um ciclo de vida complexo, ao longo do qual passam por quatro estágios bem distintos: ovo, larva (lagarta), pupa (crisálida) e adulto (imago). Pré-pupa é uma fase de transição entre larva e pupa, reconhecida por alguns autores como um estágio em separado. Em muitas espécies, o estágio adulto é relativamente curto, enquanto a duração dos estágios imaturos (ovo, larva, pupa) chega a corresponder a mais de três quartos de todo o ciclo de vida. Larvas de borboletas e mariposas são também chamadas de lagartas. Trata-se de um estágio particularmente ativo, durante o qual o inseto se alimenta e cresce em ritmo (quase sempre) acelerado. Em algumas espécies, as lagartas também estocam em tecidos do próprio corpo o alimento que será consumido mais tarde pelos adultos, que vivem pouco e não chegam a se alimentar. Em termos evolutivos, podemos dizer que as lagartas enfrentam um dilema vital: maximizar a fecundidade dos adultos, alcançando um tamanho pupal máximo, ou acelerar o desenvolvimento, antes que o alimento se deteriore ou os predadores apareçam? Para complicar ainda mais as coisas, esse dilema deve ser equacionado em um contexto ecológico heterogêneo e dinâmico, no qual as variáveis mais importantes, como a disponibilidade de recursos, estão sempre mudando. O ambiente de uma lagarta é formado por elementos que a afetam, positiva ou negativamente, incluindo componentes bióticos (competidores, parasitas, predadores etc.) e abióticos (luz solar, água, temperatura etc.). Em termos funcionais, podemos dividir os elementos que integram o ambiente de um organismo em duas categorias: recursos (elementos bióticos ou abióticos que podem ser consumidos ou explorados) e condições de vida (elementos abióticos que não são consumíveis, mas que podem afetar a aquisição ou o uso que se faz de recursos). A temperatura ambiente é um exemplo de elemento não-consumível de importância virtualmente universal. A razão para isso é que os efeitos da temperatura sobre os objetos do mundo físico estão por toda parte. Quer dizer, a temperatura muda o estado das coisas. Para os animais ectotérmicos, como uma lagarta em crescimento, temperaturas elevadas em geral resultam em melhorias nos índices de desempenho. Essa melhoria, claro, não prossegue indefinidamente. Cedo ou tarde esbarramos em uma temperatura máxima, acima da qual há um declínio brusco e acentuado nos índices de desempenho. Esses e outros efeitos da temperatura na vida dos insetos são conhecidos há bastante tempo e não surpreende que a literatura entomológica sobre o assunto seja tão rica e variada. A maioria dos estudos, porém, tem sido conduzida em países temperados ou sob condições de laboratório. Ademais, no caso específico de insetos imaturos, muitos pesquisadores estão mais interessados em encontrar respostas específicas para problemas imediatos, relacionados principalmente ao controle de pragas agrícolas e vetores de doenças, o que tem inibido o desenvolvimento de teorias mais gerais em ecologia térmica de insetos [4]. Para visualizar a relação entre temperatura e um determinado índice de desempenho, podemos começar distribuindo os dados obtidos em um gráfico formado por um par de eixos ortogonais (Figura 1). No eixo horizontal (eixo-x ou eixo das abcissas), colocamos a escala que retrata a variação na temperatura ambiente; no eixo vertical (eixo-y ou eixo das ordenadas), a escala que retrata a variação observada no índice de desempenho estudado. No fim do processo, temos uma figura, formada por uma nuvem de pontos. A idéia então é obter uma equação matemática que capture as propriedades fundamentais da figura. Essa equação costuma reduzir a nuvem original de pontos a um segmento de reta ou de algum outro tipo de curva [5]. Um exemplo desse processo pode ser visto na figura a seguir.
Figura 1. A aceleração na taxa de desenvolvimento de pupas (N = 13) de Hypothyris ninonia como uma função linear da temperatura. Nesse caso, um segmento de reta é uma representação adequada do fenômeno. Graus-dias A forma assumida pela curva que relaciona a temperatura com os índices de desempenho é descrita com precisão por modelos matemáticos relativamente sofisticados. Todavia, dentro dos limites de temperatura que os insetos normalmente experimentam no campo, esses índices, em especial o tempo de desenvolvimento e a taxa de crescimento, podem ser descritos como funções lineares da temperatura. Quando a relação é efetivamente linear, a temperatura experimentada por um organismo ectotérmico pode ser resumida por um único valor, a quantidade de graus-dias. De um ponto de vista ecológico, dois parâmetros são suficientes para caracterizar o modelo de graus-dias, a saber: (i) o limite inferior de desenvolvimento (LID), que é a temperatura abaixo da qual o desenvolvimento cessa, ou ao menos deixa de ser mensurável, e (ii) a soma das temperaturas efetivas (STE), que é o número de graus-dias (GD) acima de LID necessários para que o inseto complete alguma fase do desenvolvimento [6]. LID e STE variam de acordo com a espécie, mas também podem variar entre populações co-específicas ou até mesmo entre diferentes estágios do ciclo de vida de um inseto. Estudos sobre a acumulação de graus-dias tradicionalmente empregam dados obtidos em regimes de temperatura constante no laboratório para predizer eventos no ciclo de vida do inseto no campo. O procedimento comum nesses estudos é criar grupos de imaturos da espécie de interesse, cada um sob determinada temperatura, até a conclusão de uma fase ou de todo o ciclo de vida. Poderíamos acompanhar, por exemplo, grupos de ovos de uma espécie de inseto criados a 15, 20, 25 e 30°C, anotando o tempo que as larvas demorariam a eclodir em cada caso. Tal procedimento, no entanto, tem os seus problemas. Resultados de estudos em laboratório têm sido usados com algum sucesso para prever a ocorrência ou mesmo a abundância de populações de insetos no campo. As discrepâncias, no entanto, não são raras, pois o tempo de desenvolvimento pode diferir muito entre um regime de temperatura constante (laboratório) e um regime de temperatura variável (campo), mesmo quando a temperatura média final é a mesma [7]. Por exemplo, crescer durante 16 horas (dia) a 25°C e outras 8 horas (noite) a 18°C dentro de uma câmara de laboratório, não é a mesma coisa do que crescer sob um regime flutuante ao ar livre, mesmo quando as temperaturas médias finais são idênticas. Há, portanto, uma razoável margem de dúvida sobre a pertinência de levar para o campo os resultados obtidos em laboratório. Essas discrepâncias entre resultados obtidos em laboratório e no campo tendem a ser ainda mais acentuadas em espécies que termorregulam. Esse é o caso dos imaturos de muitos insetos que, em condições naturais, se comportam de modo a aumentar ativamente a temperatura do corpo (selecionando microhábitats mais quentes ou tomando banhos de sol, por exemplo) e, com isso, encurtar o tempo de desenvolvimento. Em linhas gerais, portanto, podemos dizer que há uma boa margem de dúvidas sobre a pertinência de extrapolar para o campo os resultados obtidos em laboratório. Vamos ver um pouco como as coisas se passam no campo. Sazonalidade reprodutiva Para ilustrar com mais detalhes a influência que a temperatura exerce sobre o desenvolvimento de insetos, vou utilizar aqui alguns dados sobre os imaturos de uma borboleta neotropical, Hypothyris ninonia daeta (Lepidoptera: Nymphalidae: Ithomidae). O crescimento das lagartas dessa borboleta tem sido objeto de estudo, tanto no campo como em laboratório [8]. A exemplo do que ocorre com outros ectotérmicos, o desempenho desses insetos é sensível à temperatura e varia ao longo do ano. Um estudo de campo descobriu, por exemplo, que a taxa de crescimento das lagartas de Hypothyris aumenta linearmente com a temperatura ambiente (dados não-publicados), a exemplo do que ocorre com a taxa de desenvolvimento das pupas (Figura 1). Na verdade, os efeitos da temperatura são notáveis: do mês mais quente para o mês mais frio, o tempo de duração de todo o estágio larval pode mais do que dobrar, ainda que a diferença na temperatura média entre os dois meses não tenha ultrapassado aos 6 graus centígrados. O padrão geral seria o seguinte: a taxa de crescimento das lagartas de Hypothyris é alta nos meses quentes de verão, principalmente em janeiro e fevereiro, diminui durante o outono, alcançando seu valor mais baixo por volta de maio e junho, quando uns poucos imaturos ainda são encontrados no campo. Por fim, nos últimos meses do inverno (agosto e setembro), já não há mais imaturos - ao que parece, por causa de uma suspensão temporária que há nas atividades reprodutivas dos adultos. Eles reaparecem na primavera, tornando-se mais numerosos a partir de novembro-dezembro, quando então o ciclo anual recomeça. O que esses resultados mostram é que a sazonalidade na temperatura ambiente pode afetar o desempenho de lagartas tropicais de modo significativo, a exemplo do que já foi descrito para diversas espécies de insetos de zonas temperadas e subárticas [9]. Para os organismos ectotérmicos, portanto, é como se todas as coisas ficassem mais lentas à medida que a temperatura ambiente diminui de modo consistente, como em geral ocorre durante o inverno. Muitos insetos talvez até pudessem continuar fazendo no inverno o que fazem no verão; todavia, como tudo se torna mais lento (no caso de insetos imaturos, por exemplo, o tempo necessário para completar o desenvolvimento tende a aumentar bastante), as chances de sobrevivência tenderiam a ser bem menores. Em outras palavras, as baixas temperaturas do inverno não representam um problema ou uma ameaça por si só, mas sim em função de certos eventos relacionados ao contexto ecológico, como a escassez de alimento ou os riscos aumentados de morte causada por predadores. Em casos assim, o comportamento mais eficiente e vantajoso para os adultos de Hypothyris talvez seja a suspensão das atividades reprodutivas durante a estação mais fria (e seca) do ano. Em meados da primavera, quando a pluviosidade aumenta e a temperatura do ar volta a subir, todo mundo poderia então retomar suas atividades plenamente e a um ritmo cada vez mais agitado, à medida que o verão se aproxima. Generalizando, embora os problemas vitais mais importantes enfrentados pelos insetos tropicais possam não ser impostos diretamente pelo ambiente físico, a sazonalidade observada em alguns de seus elementos, notadamente a temperatura do ar, cumpre um papel ecológico fundamental. Na verdade, essa talvez seja uma experiência de vida comum a quase todos nós; afinal, mesmo os moradores das grandes cidades brasileiras estão acostumados a enfrentar surtos populacionais periódicos. Ou será que alguém ainda não percebeu que moscas, pernilongos, baratas e outros tantos personagens que proliferam em hábitats urbanos tendem a escassear no inverno, para então ressurgir meses depois, quando a temperatura do ar volta a subir?
Notas
(*) Biólogo meiterer@hotmail.com, autor do livro ECOLOGIA, EVOLUÇÃO & O VALOR DAS PEQUENAS COISAS (2003).
1. As expressões ectotérmico e endotérmico não devem ser confundidas com expressões relacionadas, como é o caso de heterotérmicos (ou pecilotérmicos) e homeotérmicos. Organismos heterotérmicos são aqueles cuja temperatura interna varia mais ou menos de acordo com a temperatura ambiente. Ectotérmicos em geral também se comportam como heterotérmicos. Por sua vez, organismos homeotérmicos são aqueles cuja temperatura interna é regulada de modo a permanecer mais ou menos constante, a despeito de oscilações na temperatura ambiente. Endotérmicos geralmente se comportam como homeotérmicos, mas certos ectotérmicos também podem fazê-lo. Quando um lagarto entra e sai de cima de uma pedra na qual toma banho de sol, por exemplo, ele está regulando (de modo ectotérmico) sua temperatura interna, mantendo-a mais ou menos constante e comportando-se, portanto, como um organismo homeotérmico (ainda que apenas durante algumas horas do dia). Para detalhes, ver Schmidt-Nielsen, K. 1996. Fisiologia animal: adaptação e meio ambiente, 3a edição. SP, Santos. |
||